En una reciente cooperación con la Universidad de Utrecht, se evaluó el impacto reproductivo de DON y BEA en cerdas jóvenes y cerdas con resultados sorprendentes. Este artículo resume las principales conclusiones del estudio.
El deoxinivalenol (DON) y la beauvericina (BEA) son toxinas producidas por la especie Fusarium que pueden contaminar los alimentos y los piensos. El impacto negativo causado por el DON en los cerdos es bien conocido, y los estudios también indican que los cerdos son sensibles a la BEA. En Europa, se recomienda que las dietas para cerdos no contengan más de 900 ppb de DON, independientemente de la edad del animal; sin embargo, no existe ninguna recomendación sobre los niveles máximos de BEA en la dieta
Equilibrio redox en cerdas y cerdas jóvenes
Además de ser tóxicos para el tracto intestinal, tanto el DON como la BEA pueden perjudicar la reproducción de las hembras de cerdo, concretamente al causar daños en sus gametos, también llamados ovocitos. Estas células reproductoras mantienen constantemente un equilibrio entre la producción de especies reactivas de oxígeno (ROS) y su actividad antioxidante para garantizar un sistema reproductivo saludable. En una fase avanzada de desarrollo, estos ovocitos se encierran en un folículo y están rodeados de células somáticas (células del cúmulo) y de un líquido folicular (FF) producido por las células somáticas. La comunicación de estos compartimentos a través de señales endocrinas y el equilibrio redox mantienen la calidad de los ovocitos para su posterior fecundación. En las cerdas jóvenes prepúberes, las señales endocrinas para iniciar la pubertad permiten un crecimiento de los folículos de hasta aproximadamente 6 mm, pero no apoyan su desarrollo final. Por lo tanto, el primer celo de las cerdas jóvenes se clasifica como un celo-no-servicio. Esto es probablemente un resultado adicional de un equilibrio redox aún no eficiente. Basándonos en esta información, planteamos la hipótesis de que los oocitos de cerdas y cerdas jóvenes responderán de forma diferente a la exposición a DON y BEA durante la maduración in vitro y que el sistema redox folicular juega un papel importante en estas diferencias. Para evaluar esta hipótesis, evaluamos el efecto de DON y BEA a diferentes concentraciones en la maduración in vitro de ovocitos de cerdas y cerdas jóvenes. Además, se establecieron los niveles de antioxidantes en los ovocitos y en las células del cumulus, así como la expresión relativa del mRNA de los marcadores redox.
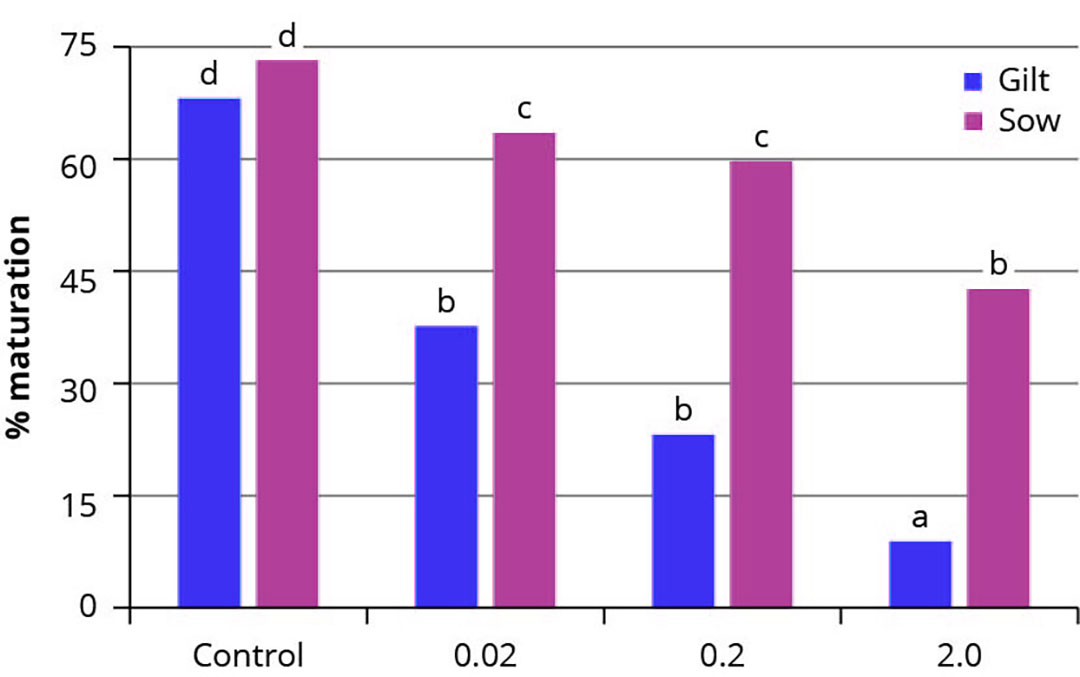
Figura 1 – Tasas medias de maduración y degeneración de ovocitos porcinos madurados in vitro en ausencia (control) o en presencia de DON a diferentes concentraciones (0,02, 0,2 o 2,0 µmol/L).
Maduración de ovocitos in vitro
Como se confirmó mediante el análisis de la expresión del ARNm, los ovocitos madurados in vitro de cerdas jóvenes tenían una menor expresión de la superóxido dismutasa 1 (SOD1) y de la glutatión sintetasa (GSS) que las cerdas. Esto indica que los ovocitos de las cerdas son más activos en la neutralización de los aniones superóxido en el citoplasma de estos gametos femeninos y que la GSS favorece el desarrollo posterior de los embriones. Los oocitos y las células del cumulus que los rodean, procedentes de cerdas y cerdas jóvenes, se evaluaron antes y después de la maduración in vitro en ausencia o presencia de DON (0,02, 0,2 o 2,0 µmol/L) o BEA (0,5, 2,5 o 5,0 µmol/L). Los resultados se detallan en las figuras 1 y 2
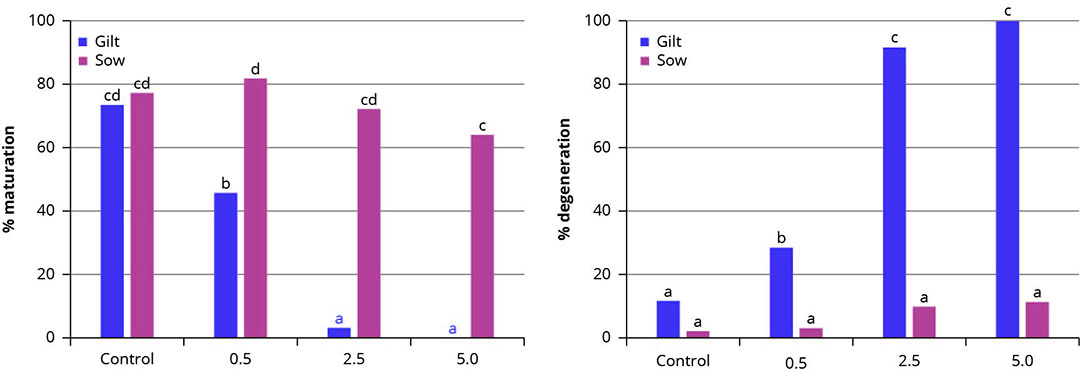
Figura 2 – Tasas medias de maduración y degeneración de ovocitos porcinos madurados in vitro en ausencia (control) o en presencia de BEA a diferentes concentraciones (0,5, 2,5 o 5,0 µmol/L).
Presencia de DON
Independientemente de la edad (cerdas o cerdas jóvenes), se observó una disminución significativa de la tasa de maduración de los ovocitos cuando éstos fueron madurados in vitro en presencia de 0,02 o 0,2 µmol/L de DON, y este impacto negativo fue aún más prominente a una concentración de 2,0 µmol/L. Además, los ovocitos de cerda fueron más sensibles a la exposición al DON que los de cerda. La tasa de degeneración osciló entre el 8,8% y el 35,4%, pero no se observó ningún efecto en la exposición al DON(Figura 1).
Efecto de la beauvericina en los ovocitos de las cerdas
Se observó una disminución significativa de la tasa de maduración de oocitos en oocitos de cerda ya en presencia de 0,5 µmol/L de BEA, y este impacto negativo fue aún más prominente en concentraciones de 2,5 o 5,0 µmol/L. Por el contrario, la BEA no afectó a la maduración de los ovocitos de cerda en las concentraciones probadas en comparación con el control. Asimismo, la tasa de degeneración de los ovocitos de cerda aumentó significativamente en presencia de BEA, independientemente de la concentración probada, mientras que BEA no alteró significativamente la degeneración de los ovocitos de cerda. Tanto para la tasa de maduración como para la de degeneración, hubo un efecto lineal significativo con el aumento de la concentración de BEA en el medio de maduración(Figura 2)
DON y BEA más tóxicos para las cerdas
En conclusión, los ovocitos de las cerdas jóvenes son más susceptibles a los efectos tóxicos del DON y la BEA en comparación con los ovocitos de las cerdas. Este efecto se observa cuando los ovocitos inmaduros comienzan su maduración. Además, el equilibrio redox en las cerdas está mejor controlado que en los ovocitos de cerdas jóvenes y está relacionado principalmente con el desequilibrio de SOD y GSH en los ovocitos. Tanto DON como BEA no afectan a la viabilidad de la célula sólo a través de la estimulación del estrés oxidativo, pero su impacto negativo se potenciará en las células que no pueden eliminar adecuadamente los productos metabólicos como ROS. Los niveles de exposición probados de DON en el presente estudio son como los detectados en el nivel de plasma porcino en los cerdos expuestos crónicamente a DON que pueden llegar a 0,09 µmol/L. La detección de BEA en muestras biológicas es un proceso complejo porque esta micotoxina está presente en la dieta de estos animales en concentraciones mucho más bajas que el DON y se metaboliza rápidamente en una serie de derivados aún no caracterizados. Sin embargo, en el caso de ambas micotoxinas, es conveniente prestar especial atención a la contaminación de la dieta de las cerdas jóvenes.
El estudio se realizó bajo la supervisión del Dr. Bernard Roelen. Eric J. Schoevers
†
falleció antes de la publicación de este trabajo.